真核生物の起源は生物学における大きな未解決問題です。近年、環境ゲノムの解析によって真核生物特有と思われてきた遺伝子を多数有するアスガルド上門アーキア(アスガルド古細菌)と呼ばれる微生物群が発見され、真核生物の直接の祖先生物の可能性が示唆されています。アスガルド古細菌で見つかった真核生物様の遺伝子をタンパク質レベルで分析すると、実際に真核生物でみられるような形状や機能が保存されていることが徐々に明らかになりつつあります。
東京工業大学 地球生命研究所 (ELSI)のCaner Akıl研究員らは、これまでにアスガルド古細菌から多数のアクチン制御タンパク質を発見しており、それらの最新の知見を本論文にまとめました (Akıl et al., 2020, Curr. Opin. Cell Biol)。アクチン動態における機能を調べることで、古細菌から真核生物への進化において重要な機能を果たしたとされるアクチン細胞骨格の制御系の起源に迫ることができると研究チームは考えています。
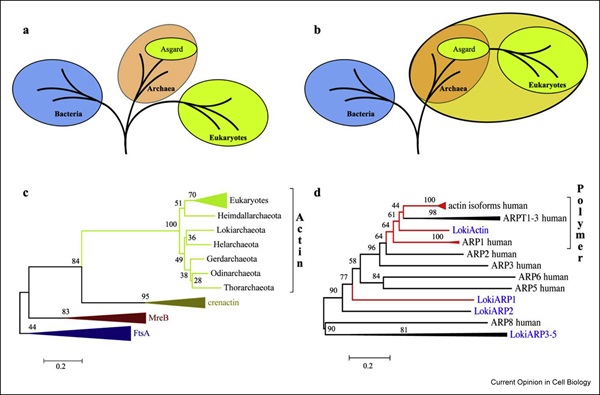
真核細胞の特徴は、複雑に制御されたアクチン細胞骨格の存在です。アクチン細胞骨格の機能は、主に細胞膜の修復や細胞そのものの動きに深く関わっています。面白いことに、近年、アスガルド古細菌は、真核生物と共通の祖先に由来する原始的なアクチン制御タンパク質を保有することがわかってきました。
Akıl研究員(研究当時シンガポール国立大学の博士課程)と岡山大学のRobert C. Robinson教授は、生化学的かつ生物物理学的観点から真核生物が持つアクチン調整タンパク質であるプロフィリンに似たタンパク質をアスガルド古細菌が持っていることを初めて示唆しました。プロフィリンは、真核生物の細胞骨格を調整するための主要な役割を果たしています。真核生物の場合、ポリプロリンの結合部位でフィラメント核生成部分に対してアクチンとプロフィリンの複合体が供給されますが、アスガルド古細菌のプロフィリンは、このような結合部位を持っていません。このようなアクチンフィラメント核生成タンパク質は、アスガルドのゲノムではまだ発見されていません。(Akıl & Robinson, 2018, Nature)。
一方でAkil研究員らは別のアクチン調節タンパク質である真核生物様ゲルソリンをアスガルド古細菌のトール門から発見しました。アスガルド古細菌におけるゲルソリンは、遺伝子重複によって生じたと考えられるコフィリン/ゲルソリンドメインの繰り返しパターンを有しており、真核生物で見られるようなマルチドメイン型ゲルソリンの原型であると考えられます。
また面白いことに、アスガルド古細菌の持つゲルソリンのタンパク質の多くの構造は門によって異なることから、アスガルド上門の進化の過程で高度なアクチン調節能を獲得したと考えられます。実際に真核生物では実に多くのアクチン制御タンパク質が知られており、それはアクチンフィラメントの重合や分解、キャッピング、修飾、核形成、束化など、その機能は多岐にわたります。それらに比べれば、アスガルド古細菌におけるアクチン制御タンパク質は実にシンプルですが、それでも動的なフィラメント形成やその構造制御は真核生物のタンパク質の機能と酷似していることをAkıl研究員らは明らかにしました。これは実際にアスガルド古細菌から真核生物への進化を示す確固たる証拠の一つといえるでしょう (Akıl et al., 2020, PNAS)。
今後も、アスガルド古細菌における機能未知タンパク質群から、アクチン動態を制御する因子が新たに発見されることでしょう。そして最近その特徴的な細胞形状が明らかとなり大きな話題となったMK-D1株のように、アスガルド古細菌のそれぞれの門ごとに保有しているタンパク質の種類と形状の関連性が明らかとなることで、真核生物の初期のアクチン動態に関する重要な知見が得られると研究チームは期待しています。
| 掲載誌 | Current Opinion in Cell Biology |
| 論文タイトル | Mythical origins of the actin cytoskeleton |
| 著者 | Caner Akıl1,2,a, Yoshihito Kitaoku3,a Linh T. Tran3, David Liebl4, Han Choe5, Duangkamon Muangsaen6, Wipa Suginta6, Albert Schulte6 & Robert C. Robinson1,3,6* |
| 所属 | 1 Institute of Molecular and Cell Biology, A*STAR (Agency for Science, Technology and Research), Biopolis, 138673, Singapore 2 Tokyo Institute of Technology, Earth-Life Science Institute (ELSI), Tokyo, 152-8551, Japan 3 Research Institute for Interdisciplinary Science (RIIS), Okayama University, Okayama 700-8530, Japan 4 A*STAR Microscopy Platform, Research Support Center, A*STAR, Biopolis, 138673, Singapore. 5 Department of Physiology, University of Ulsan College of Medicine, Asan Medical Center, Seoul, 05505, Korea 6 School of Biomolecular Science and Engineering (BSE), Vidyasirimedhi Institute of Science and Technology (VISTEC), Rayong 21210 Thailand |
| DOI | 10.1016/j.ceb.2020.08.011 |
| 出版日 | 2020年10月10日 |