Using a cell-free translation system known as the PURE system, a team of researchers along with ELSI members have optimized a protein selection tool known as the mRNA/cDNA display method. This method, in general, enables a screening of over 1012 random protein sequences in a single experiment based on versatile function. To validate the presented method's performance, the team targeted a commercially available antibody that strongly binds to a specific peptide sequence known as FLAG. As a result, high-throughput sequencing data revealed an enriched known consensus FLAG sequence with additional unreported features. Hence, this approach will be helpful to explore the sequence and functional space of diverse polypeptides.
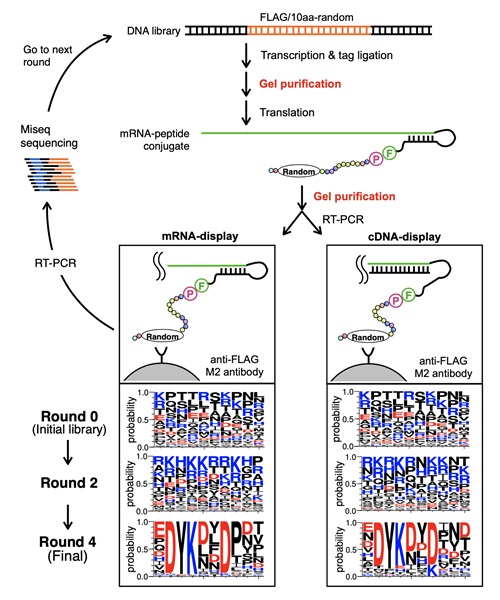
Cell-free transcription and translation systems have become an essential tool for in vitro protein expression mainly due to their flexibility in the reaction conditions and minimizing the limitations of working with actual living cells. The PURE system offers a contaminant-free solution by only using the purified components required for protein synthesis. The researchers have applied the PURE system to the mRNA display method and introduced two additional gel purification steps to maximize the products' purity. These improvements resulted in the improved formation of chemically conjugated mRNA (genotype) and its coding protein (phenotype). A cDNA display requires an additional step to mRNA display, making a complementary cDNA strand of the mRNA to prevent secondary structure formation and increase its stability. In general, these display methods can screen a vast number of protein molecules (up to 1013) in a single experiment. They targeted commercially available anti-FLAG antibodies for evaluating the newly refined display method's performance due to the nature of its known short consensus binding motif (DYKXXD). Starting from approximately 1.7 x 10^12 random peptide sequences, the team performed a total of four selection rounds for mRNA and cDNA display (Figure 1).

A round-by-round high-throughput sequencing revealed clear enrichment of the FLAG epitope consensus motif DYK(D/L/N)(L/Y/D/N/F)D for both display methods, which required minimum three rounds of selection for the motifs to become prominent (Figure 2). Overall, both display methods resulted in similar performance with slightly higher enrichment for mRNA display (Figure 3).
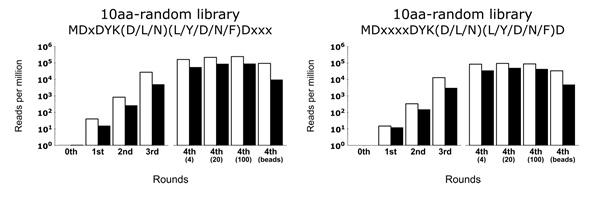
Whereas, a control experiment eliminating the gel purification step resulted in poor conjugate product formation and led to an increase of sequences with incorrect lengths. It indicates that conjugate purity contributes to the rapid enrichment of correct binding sequences. Under such messy conditions, the cDNA display has an advantage over the mRNA display, minimizing the non-specific interaction by masking the mRNA region with cDNA. The newly refined in vitro selection tool will allow researchers to explore diverse protein sequence space towards understanding primitive protein functions relevant to the origin of life study as well as developing de novo protein aptamers for future biomolecular tools and peptide drug developments.
| Journal | Biotechnology and Bioengineering |
| Tile of the paper | PURE mRNA display and cDNA display provide rapid detection of core epitope motif via high‐throughput sequencing |
| Authors | Sabrina Galiñanes Reyes1,2,3, Yutetsu Kuruma1,2,4, Mai Fujimi1, Masako Yamazaki5, Sumie Eto1,5, Shota Nishikawa1,6, Satoshi Tamaki5, Asaki Kobayashi7, Ryo Mizuuchi4,8, Lynn Rothschild9, Mark Ditzler9, Kosuke Fujishima1,10,* |
| Affiliations |
1Earth‐Life Science Institute, Tokyo Institute of Technology, Meguro‐ku, Tokyo, Japan 2Extra‐cutting‐edge Science and Technology Avant‐garde Research Program, Japan Agency for Marine‐Earth Science and Technology, Kanagawa, Japan 3James Watt School of Engineering, The University of Glasgow, Glasgow, UK 4JST, PRESTO, Saitama, Japan 5MOLCURE Inc., Shinagawa, Tokyo, Japan 6School of Life Science and Technology, Tokyo Institute of Technology, Tokyo, Japan 7SABNP, Univ Evry, INSERM U1204, Université Paris‐Saclay, Evry, France 8Komaba Institute for Science, The University of Tokyo, Meguro‐ku, Tokyo, Japan 9Center for the Emergence of Life, NASA Ames Research Center, Moffett Field, California, USA 10Graduate School of Media and Governance, Keio University, Fujisawa, Japan |
| DOI | 10.1002/bit.27696 |
| Online published date | 27 January, 2021 |